Effects of Polymorphisms of Innate Immunity Genes and Environmental Factors on the Risk of Noncardia Gastric Cancer
Article information
Abstract
Purpose
Increasing evidence suggests that polymorphisms in innate immunity genes are associated with Helicobacter pylori-induced inflammation and may influence susceptibility in developing noncardia gastric cancer. Therefore, we investigate the effect of polymorphisms of innate immunity genes and interactions with environmental factors in the Korean population.
Materials and Methods
We genotyped four polymorphisms of TLR2 (rs1898830), TLR4 (rs10983755 and rs10759932), and CD14 (rs2569190) in a case-control study of 487 noncardia gastric cancer patients and 487 sex- and age-matched healthy controls. Polytomous logistic regression models were used to detect the effects of genetic polymorphisms and environmental factors, which were stratified by the histological type of gastric cancer.
Results
TLR4 rs10983755 A carriers were found to have higher risk of intestinal-type noncarida gastric cancer than G homozygotes (odds ratio [OR], 1.41; 95% confidence interval [CI], 1.01 to 1.97), but other genetic variants showed no association with the risk of noncardia gastric cancer. Among H. pylori-positive participants, smokers carrying TLR4 rs10983755 A had a higher risk of intestinal-type gastric cancer than nonsmoking TLR4 rs10983755 G homozygotes (OR, 4.28; 95% CI, 2.12 to 8.64). In addition, compared with tap water, other drinking water sources during childhood were found to be associated with the elevated risk of intestinal-type gastric cancer, and these associations were slightly stronger among TLR4 rs10983755 A carriers.
Conclusion
The genetic polymorphisms of innate immunity genes are associated with the development of intestinal-type noncardia gastric cancer and these associations may differ in accordance to an exposure to certain environmental factors.
Introduction
Helicobacter pylori is the most important risk factor for gastric cancer, and its association with the innate immune response, especially toll-like receptors (TLRs), is important in the inflammatory response of gastric mucosa [1,2]. TLRs, particularly TLR2 and TLR4, play a role in the recognition of H. pylori and activate multiple intracellular signaling pathways that culminate in the nuclear factor-κB activation and subsequent production of inflammatory and immune effectors [1]. TLR4 recognizes lipopolysaccharide (LPS), a major cell wall constituent of gram-negative bacteria, for which CD14 is a receptor [1]. Variations in TLR genes have been proposed to alter the ligand binding and balance between pro- and anti-inflammatory cytokines, thereby modulating the risk of chronic inflammation and cancer [2]. Additionally, environmental factors may also affect an individual's susceptibility to the long-term outcome of H. pylori infection and gastric cancer development by interacting with the genetic factors [3].
To date, several studies have investigated the role of innate immunity gene polymorphisms on gastric carcinogenesis [4-6]. However, which functional polymorphisms of innate immunity gene account for H. pylori-related gastric malignancies remains unknown. Additionally, strong ethnic differences exist in the distribution of those polymorphisms [2]. Therefore, this study aims to elucidate the potential association of innate immunity genes such as TLR2, TLR4, and CD14 with the risk of noncardia gastric malignancies, which are more related to H. pylori infection and chronic inflammation than cardia gastric cancer [7], in the Korean population. Additionally, we examine the differential effects of these genes on gastric cancer risk according to other risk factors, including age, tobacco smoking, alcohol consumption, and drinking water source during childhood.
Materials and Methods
1. Study population
This study was a hospital-based case-control study consisting of 797 patients with noncardia gastric cancer, who were recruited from the National Cancer Center Hospital in Goyang between 2003 and 2007. Histologically confirmed adenocarcinoma patients were eligible for participation in this study; other types of neoplasms in the stomach, including gastric lymphoma, gastrointestinal stromal tumors, carcinoid tumors, or adenomas, were excluded. The controls consisted of 930 healthy participants without significant gastrointestinal symptoms who were recruited in 2007 from the participants who underwent upper endoscopy for gastric cancer screening of the National Cancer Screening Program in Korea. All cases and controls were of Korean descent. We excluded individuals with a history of other malignant neoplasms, with a history of H. pylori eradication therapy, or without H. pylori evaluation and genotyping information.
Cases and controls were frequency-matched by age (within 5 years) and sex. The final analysis included 487 cases and 487 controls. Noncardia gastric cancer patients were classified into intestinal or diffuse-type based on the histopathology, according to the Lauren classification [8]. The clinical information and genetic materials were obtained with an approval from the Institutional Review Board of the National Cancer Center, and written informed consent was obtained from each participant.
2. Data collection
All questionnaire data and blood samples were collected at the initial recruitment of both the cases and controls. Participants were asked to complete a self-administered questionnaire regarding their sociodemographic characteristics (e.g., age, family history of gastric cancer, income, and education), lifestyles (e.g., smoking habits and alcohol intake), and personal medical history. H. pylori infection was evaluated using a rapid urease test and a histological evaluation. The rapid urease test (Pronto Dry, Medical Instruments Co., Solothurn, Switzerland) was performed in accordance to the manufacturer's instructions. For the histological evaluation of H. pylori, biopsy specimens were obtained from three different portions of the stomach-the gastric antrum lesser curvature, the corpus lesser curvature, and the corpus greater curvature-to increase the sensitivity of the procedure. Two pieces of biopsy specimens were obtained from each site, and Wright-Giemsa staining was routinely applied on the biopsy samples to evaluate the presence of H. pylori.
3. Genotyping
Genotyping of genetic variants of TLR2 (rs1898830), TLR4 (rs10759932 and rs10983755), and CD14 (rs2569190) was performed and interpreted by researchers blinded to case/control status. The related information of these single nucleotide polymorphisms (SNPs) is listed in Appendix 1. We prepared genomic DNA from peripheral blood samples using an automatic DNA extraction system (BioRobot M48 Workstation, Qiagen, Inc., Valencia, CA) according to the manufacturer's instructions, and determined the purity and concentration of isolated DNA using PicoGreen (Molecular Probes, Inc., Eugene, OR). A total of 10 ng per sample was used for genotype analysis, and genotyping for four SNPs was achieved using the iPLEX Gold assay (Sequenom, San Diego, CA), which is based on matrix-assisted laser desorption/ionization time-of-flight spectrometry according to the manufacturer's protocol. The resulting genotype data were obtained using Typer v4.0 (Sequenom). Polymerase chain reaction (PCR) primer sequences are shown in Appendix 1. PCR amplification started at 94℃ for 15 minutes, followed by 45 cycles of 94℃ for 20 seconds, 50℃ for 30 seconds, and 72℃ for 1 minute, with final extension of 72℃ for 3 minutes. After dephosphorylation of PCR products, PCR products were inputted to the allele-specific single base primer extension reactions. For quality control, subsets of samples were run as duplicates, and 5% of the wells in a 384-well plate were used as negative controls to test for contamination. For all results, genotype clusters were examined manually for their fitness. The genotyping call rates were 0.99 for TLR4 rs10759932 and 0.96 for TLR4 rs10983755, TLR2 rs1998830 and CD14 rs2569190.
4. Statistical analysis
Chi-square (χ2) test and t-tests were used to compare the demographic characteristics and environmental factors in the cases and controls. The Hardy-Weinberg equilibrium test was conducted for each SNP among the controls. The association between genetic variants of TLR2, TLR4, and CD14 and gastric cancer was estimated by polytomous logistic regression, stratified by histological type of gastric cancer (intestinal/diffuse-type). In multivariate analyses, data were adjusted for age, sex, H. pylori infection, smoking, alcohol consumption, education, and drinking water source during childhood, and were expressed as odds ratios (ORs) with 95% confidence intervals (CIs). Multiple comparisons were conducted by both the false discovery rate (FDR) at the 5% level and Bonferroni correction. We also conducted a haplotype analysis for two SNPs of TLR4.
The biological interaction between the two polymorphisms was estimated and its significance was assessed using the likelihood ratio test, comparing the model with the interaction term with one containing only the main effects, if necessary. To assess whether the effect of the studied polymorphisms was modified by other risk factors (e.g., age, smoking, alcohol consumption, and drinking water source during childhood), we performed a stratified analysis. Subjects were dichotomized according to age (<55 years vs. ≥55 years), smoking status (smokers vs. nonsmokers), and alcohol consumption status (drinkers vs. nondrinkers). We also tested the role of the source of drinking water (e.g., tap water, ground water, well water, and natural/mineral water) during childhood. All data were stratified by H. pylori infection status (H. pylori-positive vs. H. pylori-negative). All statistical analyses were performed using SAS ver. 9.1 (SAS Institute Inc., Cary, NC), and two-sided p-values of less than 0.05 were regarded as statistically significant.
Results
The characteristics of the study participants are shown in Table 1. No differences were found in the distribution of age, sex, family history of gastric cancer, or alcohol consumption between the cases and controls; however, the cases were more likely to have H. pylori infection (p<0.001), more likely to smoke (p<0.001), and less likely to drink tap water and more likely to drink well water during childhood (p<0.001) than the controls. In addition, the cases had less education (p<0.001) than the controls.
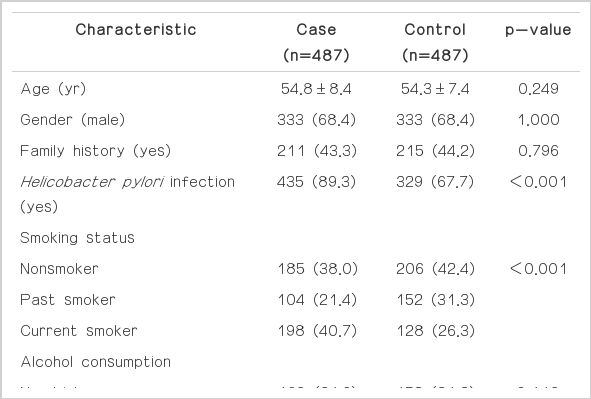
Characteristics of study participants
The genotype frequencies of all tested polymorphisms in the controls did not deviate from the Hardy-Weinberg equilibrium (data not shown). Regarding genetic frequencies, no significant differences were found between the cases and controls in the frequencies of TLR4 rs10759932, TLR2 rs1898830, and CD14 rs2569130, except for TLR4 rs10983755. TLR4 rs10983755 A homozygotes (OR, 2.08; 95% CI, 1.09 to 4.00) and A allele carriers (OR, 1.41; 95% CI, 1.01 to 1.97) were found to have a higher risk of intestinal-type noncardia gastric cancer than G homozygotes. Other genetic variants showed no association with noncardia gastric cancer risk. However, none of the tested values were significant after testing with both the FDR at the 5% level and Bonferroni correction (Table 2). In addition, we examined the linkage disequilbrium (LD) of the two polymorphisms (rs10759932 and rs10983755) in TLR4 and fount that they are in strong LD (r2=0.88, D'=0.99). We also conducted a haplotype analysis using two SNPs of TLR4. Haplotype '2-2' showed a slightly increased risk of intestinal-type gastric cancer (OR, 1.37; 95% CI, 1.05 to 1.80) (Table 2).
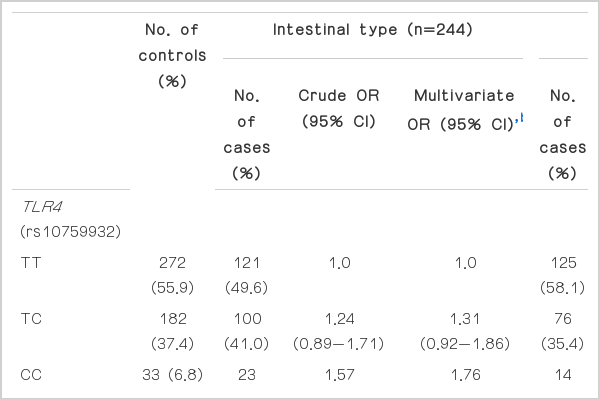
Association between genetic variants of TLR4, TLR2, and CD14 and the risk of noncardia gastric cancer, stratified by histological typea)
To assess the combined effects of tested SNPs on gastric cancer susceptibility, we investigated the association between TLR4 rs10983755 and the risk of gastric cancer, in accordance to the genotypes of TLR2 rs1898830 and CD14 rs2569190 (Table 3). The association between TLR4 rs10983755 genotype and risk of gastric cancer did not differ by TLR2 rs1898830 genotype or CD14 rs2569190 genotype. A haplotype analysis, using two SNPs of TLR4, showed a similar finding (Appendix 2).

Association between TLR4 rs10983755 and the risk of noncardia gastric cancer according to TLR2 rs1898830 and CD14 rs2569190, stratified by histological typea)
To investigate the role of other known risk factors in gastric cancer, we categorized participants according to their H. pylori infection status, and then examined the association between TLR4 genotype and the risk of gastric cancer via stratification by other risk factors (Table 4). Among H. pylori-infected participants, older people (≥55 years old) seemed to have a decreased risk for diffuse-type gastric cancer, but no difference was observed by TLR4 rs10983755 genotype. Smoking increased the risk of intestinal-type gastric cancer, with a slightly stronger association among TLR4 rs10983755 A carriers. Compared with nonsmoking TLR4 rs10983755 G homozygotes, smokers with TLR4 rs10983755 A carriers showed a significantly increased risk of intestinal-type gastric cancer (OR, 4.28; 95% CI, 2.12 to 8.64). Alcohol consumption was not associated with gastric cancer by their genetic variants and histological type. Concerning drinking water source during childhood, participants who used other sources of drinking water had a higher risk of intestinal-type gastric cancer than those who used tap water, and the association was slightly stronger among TLR4 rs10983755 A carriers. Compared with TLR4 rs10983755 G homozygotes who drank tap water during childhood, TLR4 rs10983755 G carriers had a higher risk of intestinal-type gastric cancer if ground water (OR, 2.86; 95% CI, 1.39 to 5.89), well water (OR, 2.16; 95% CI, 1.03 to 4.51), and natural/mineral water (OR, 3.45, 95% CI, 1.24 to 9.57) was their primary drinking water source during childhood. Haplotype analyses, using two SNPs of TLR4, did not present any differential associations by smoking status or drinking water source (Appendix 3).

Association between TLR4 rs10983755 and gastric cancer risk according to age, smoking, alcohol consumption status, and drinking water source during childhood among 764 Helicobacter pylori positive participants, stratified by histological typea)
Discussion
Emerging evidence has suggested that genetic polymorphisms in pathogen recognition receptors, such as TLRs, are associated with gastrointestinal cancer [4,9]. TLR4 Asp299Gly (rs4986790) and Thr399Ile (rs4986791) are the most intensively investigated polymorphisms; however, studies in different populations have reported either a positive association with gastric carcinoma [4,9] or no association [5]; these TLR polymorphisms are almost never found in the Asian populations [10,11]. Therefore, we investigated two polymorphisms of the promoter region of TLR4, located at -2,081 and -1,607 bp, and found a potential role of TLR4 rs10983755 in the risk of noncardia gastric cancer, although the significant association disappeared after multiple comparisons. To our knowledge, no study has examined the role of these polymorphisms in gastric cancer risk; however, some studies have reported their associations with other cancers. Cheng et al. [12] found that TLR4 rs10759932 C homozygotes have a 4-fold greater risk of prostate cancer than the wild-type homozygotes. However, Chen et al. [13] reported that men carrying the C allele of TLR4 rs10759932 have a lower risk of prostate cancer. Several studies have investigated the role of the CD14 rs2569190 polymorphism, but reported inconsistent findings [10,14]. No apparent association with H. pylori-related gastric malignancy was observed in the Taiwanese Chinese population [10], but a lower risk of intestinal-type gastric cancer in CD14 rs2569190 T allele carriers was found in the Japanese population [14]. Only a few studies have investigated the role of TLR2 polymorphisms in gastric carcinogenesis, but they reached no conclusions [1,6]. In the present study, no associations were confirmed for polymorphisms in CD14 and TLR2.
The mechanism of the action of innate immunity genes in gastric carcinogenesis remains unclear, but chronic inflammation due to related polymorphisms seems to play a key role [2,15]. Evidence has indicated that TLR4 promoter sequence variation may result in reduced TLR4 expression [16]. Ragnarsdottir et al. [16] reported that TLR4 promoter sequence variants reduced TLR4 expression in response to the infection in the Swedish population. Among the investigated SNPs, TLR4 rs10983755 G/A presented a change in promoter activity after the infection; it disrupted a potential binding site for N-Myc and modified the luciferase expression levels. It has been suggested that individuals with TLR4 polymorphisms are hyporesponsive to LPS, which allows H. pylori to avoid an attack from the host immune system and to persist in gastric mucosa, leading to chronic inflammation [9,15]. Fukata et al. [17] found that TLR4 -deficient mice have defective intestinal mucosal repair in response to chemically induced mucosal damage. Additionally, genetic polymorphisms polymorphisms in innate immunity genes may dysregulate TLR signaling, and thus, contribute to an unbalanced ratio between pro- and anti-inflammatory cytokines [18], causing chronic inflammation and gastric cancer. Higgins et al. [19] also demonstrated that TLR-4 defective mice have more severe infection, which was related to the reduced innate and Ag-specific interleukin-10 production and to the enhanced inflammatory cytokine production. Measuring the amount of TLR4 expression (DNA, RNA, and protein levels) and functional consequences of TLR4 polymorphisms may help to elucidate the underlying mechanism [2].
The effect of a genetic trait on the process of gastric carcinogenesis could be modified by several factors that modulate the innate immune response and cytokine production [1,20]. H. pylori infection status may affect the role of genetic or environmental factors on gastric carcinogenesis [20], and we observed the modifying effect of some risk factors only among H. pylori-infected subjects. We found an increased risk of intestinal gastric cancer among smokers, with a stronger association among TLR4 rs10983755 variant allele carriers. Smoking is known to elevate the risk of gastric cancer, synergistically with H. pylori infection [3]. Smoking seems to decrease TLR4 expression, which is already affected by genetic variants of TLR4. Chen et al. [21] reported that exposure of alveolar macrophages to tobacco smoke induces a hyporesponsive state, possibly due to inhibited TLR2/TLR4 expression, resulting in suppressed expression of proinflammatory mediators. Additionally, we found that risk of gastric cancer differed according to sources of drinking water. Compared with drinking tap water during childhood, other water sources seem to increase the risk of intestinal-type gastric cancer, and this association was more apparent among individuals with variants alleles of TLR4 in this study. H. pylori is known to be primarily acquired in childhood and the risk of infection declines rapidly thereafter [22]. Childhood H. pylori infection can occur through the consumption of H. pylori-contaminated water [23]. Individuals with TLR4 polymorphism appear to have greater susceptibility to H. pylori infection at childhood, which can result in lifetime persistent infections [20].
Our study presented differential results according to two main histological types (diffuse and intestinal), which have different clinico-pathological characteristics [8]. The intestinal-type of gastric cancer is more frequently found in old men, while the diffuse-type of gastric cancer is relatively frequent in young women [8,24]. Different precancerous lesions probably exist for these two types; the intestinal-type of gastric cancer is thought to develop after a long lead-time and arise through a multistep process from chronic gastritis, atrophy, intestinal metaplasia, and dysplasia to carcinoma [25]. However, the diffuse-type is known to develop following a chronic inflammation without passing through the intermediate steps [7]. Hold et al. [4] reported that functional TLR4 polymorphism, which may induce LPS-hyporesponsiveness, is associated with the development of precancerous abnormalities within stomach and the increased risk of noncardia gastric carcinoma. Additionally, environmental factors, such as cigarette smoking and H. pylori infection, were found to have a stronger association with the risk of intestinal-type cancer compared to the diffuse-type cancer [3]. Cigarette smoking and H. pylori infection are also known to cause precancerous lesions, such as intestinal metaplasia [25]. The association of the TLR4 polymorphisms, smoking, and H. pylori infection with both gastric carcinoma and its precursor lesions implies that it is relevant to the entire multistage process of gastric carcinogenesis [19]. Therefore, it is reasonable to conclude that these factors are closely associated with intestinal-type cancer [3].
The present study evaluated the effects of polymorphisms of several innate immunity genes on gastric cancer risk and the effect of their combination with other risk factors in a homogenous ethnic group. However, several limitations should be considered in the interpretation of our results. First, the current study has the limitations inherited in a hospital-based case-control study (e.g., selection and recall biases), which may affect the results, and inflammation-related processes among the controls could bias the results. Second, the influence of H. pylori infection on gastric cancer could be underestimated due to spontaneous clearance of the bacteria after histological changes in the stomach and the lack of information regarding H. pylori strains. Third, we dichotomized our data in accordance to smoke exposure and alcohol consumption by combining past smokers and current smokers, as well as past drinkers and current drinkers; thus, the associations may not reflect the amount and duration of exposure, which may affect the risk of gastric cancer. Fourth, along with the studied factors, we must consider several residual confounding factors, such as other genes and environmental factors (e.g., dietary habits) that may affect the pathogenesis of gastric cancer. Finally, the relatively small sample size may hinder exploration of the gene-gene and gene-environment interactions, particularly in stratified analyses. Therefore, near significant interactions or even the null results should be interpreted carefully. In addition, given the multiple comparison, the significant results for TLR4 rs10983755 could be a chance finding. Further larger studies are required to validate the findings from the current study.
Conclusion
In this study, we identified the possible effects of innate immunity polymorphisms on gastric cancer and their interactions with environmental factors. Assessment of host-environment interactions may provide clues to allow more accurate prediction of the outcomes of gastric inflammation and appropriate treatment strategies. Further large, well-designed studies are necessary to elucidate additional gene-gene and gene-environment interactions in gastric cancer susceptibility.
Acknowledgments
This study was supported by the National Cancer Center, Korea (grant number: 1010190 and 1110300).
Notes
Conflict of interest relevant to this article was not reported.
Appendices
Appendix 1
SNPs information and primer sequences
| Gene | SNP ID | Allele | Chr band | SNP position/Function | Primer sequence |
|---|---|---|---|---|---|
| TLR4 | rs10983755 | A/G | 9q33.1 | ‒1607 | F: ACGTTGGATGCAGTCCACCACAAAATGGTC |
| 5ʹ near gene | R: ACGTTGGATGCTGTAATATCCTCCAAGCAC | ||||
| E: AGCTTGGTTTTTGACAC | |||||
| TLR4 | rs10759932 | C/T | 9q33.1 | ‒2081 | F: ACGTTGGATGAAATGCAAGCTTCTGCTATG |
| 5ʹ near gene | R: ACGTTGGATGCAGGAGTTCTCATTTTTTCAC | ||||
| E: ACAGACCAGAAAGTAATAATAAG | |||||
| TLR2 | rs1898830 | A/G | 4q31.3 | 8013 | F: ACGTTGGATGCCCCTATTTTCTAGCACATT |
| Intron | R: ACGTTGGATGCCTTAAAAACTGGAAAAGGA | ||||
| E: CTTATATTATTATTTCCCCTGTTC | |||||
| CD14 | rs2569190 | C/T | 5q31.3 | ‒260 | F: ACGTTGGATGGAGACACAGAACCCTAGATG |
| 5ʹ UTR | R: ACGTTGGATGGCAATGAAGGATGTTTCAGG | ||||
| E: AGAATCCTTCCTGTTACGG |
SNP, single nucleotide polymorphisms; F, forward; R, reverse; E, extension.
Appendix 2
The association between TLR4 haplotype and gastric cancer according to TLR2 rs1898830 and CD14 rs2569190a)
| TLR4 haplotypeb) | Intestinal type |
Diffuse type |
||||||
|---|---|---|---|---|---|---|---|---|
| Control (n=487) | Case (n=487) | Crude OR (95% CI) | Multivariate (95% CI)c) | Case (n=487) | Crude OR (95% CI) | Multivariate (95% CI)c) | ||
| TLR2 (rs1898830) | ||||||||
| AA | 1-1 | 195 (72.2) | 89 (64.5) | 1.0 | 1.0 | 83 (75.5) | 1.0 | 1.0 |
| 1-2 or 2-1 | 6 (2.2) | 1 (0.72) | 0.37 (0.04-3.08) | 0.38 (0.04-3.54) | 4 (3.6) | 1.57 (0.43-5.70) | 2.04 (0.40-10.42) | |
| 2-2 | 69 (25.6) | 48 (34.8) | 1.52 (0.98-2.38) | 1.81 (1.11-2.97) | 23 (20.9) | 0.78 (0.46-1.34) | 1.07 (0.60-1.94) | |
| AG+GG | 1-1 | 529 (75.1) | 251 (71.7) | 1.0 | 1.0 | 242 (75.6) | 1.0 | 1.0 |
| 1-2 or 2-1 | 16 (2.3) | 9 (2.6) | 1.19 (0.52-2.72) | 1.27 (0.53-3.05) | 7 (2.2) | 0.96 (0.39-2.36) | 1.09 (0.42-2.86) | |
| 2-2 | 159 (22.6) | 90 (25.7) | 1.19 (0.89-1.61) | 1.24 (0.90-1.72) | 71 (22.2) | 0.98 (0.71-1.34) | 0.95 (0.68-1.34) | |
| CD14 (rs2569190) | ||||||||
| TT | 1-1 | 244 (73.1) | 107 (62.2) | 1.0 | 1.0 | 105 (71.9) | 1.0 | 1.0 |
| 1-2 or 2-1 | 6 (1.8) | 4 (2.3) | 1.52 (0.42-5.50) | 1.58 (0.42-5.91) | 6 (4.1) | 2.32 (0.73-7.37) | 2.18 (0.61-7.73) | |
| 2-2 | 84 (25.2) | 61 (35.5) | 1.66 (1.11-2.47) | 1.80 (1.17-2.77) | 35 (24.0) | 0.97 (0.61-1.53) | 1.01 (0.62-1.65) | |
| TC+CC | 1-1 | 480 (75.0) | 233 (73.7) | 1.0 | 1.0 | 220 (77.5) | 1.0 | 1.0 |
| 1-2 or 2-1 | 16 (2.5) | 6 (1.9) | 0.77 (0.30-2.00) | 0.81 (0.29-2.25) | 5 (1.8) | 0.68 (0.25-1.89) | 0.87 (0.29-2.66) | |
| 2-2 | 144 (22.5) | 77 (24.4) | 1.10 (0.80-1.51) | 1.17 (0.82-1.66) | 59 (20.8) | 0.89 (0.64-1.26) | 0.89 (0.62-1.29) | |
OR, odds ratio; CI, confidence interval. a)Polytomous logistic regression was used to assess the association by histological type of gastric cancer, b)Composed of 2 polymorphisms of TLR4 rs10759932 and rs10983755; "1" indicates the major allele of each polymorphism and "2" indicates the minor allele of each polymorphism, c)Adjusted for age, sex, Helicobacter pylori infection, smoking, alcohol consumption, education, and drinking water source during childhood.*p<0.05.
Appendix 3
The association between TLR4 haplotype and gastric cancer according to age, smoking, alcohol consumption status, and drinking water source durnig childhood among 764 Helicobacter pylori positive participants, stratified by histological typea)
| TLR4 haplotypeb) | Intestinal type |
Diffuse type |
||||||
|---|---|---|---|---|---|---|---|---|
| Control (n=320) | Case (n=170) | Crude OR (95% CI) | Multivariate (95% CI)c) | Case (n=236) | Crude OR (95% CI) | Multivariate (95% CI)c) | ||
| Age (yr) | ||||||||
| <55 | 1-1 | 241 (75.3) | 111 (65.3) | 1.0 | 1.0 | 178 (75.4) | 1.0 | 1.0 |
| 1-2 or 2-1 | 7 (2.2) | 6 (3.5) | 1.86 (0.61-5.67) | 1.89 (0.59-5.99) | 6 (2.5) | 1.16 (0.38-3.51) | 1.34 (0.42-4.28) | |
| 2-2 | 72 (22.5) | 53 (31.2) | 1.60 (1.05-2.43) | 1.66 (1.06-2.58) | 52 (22.0) | 0.98 (0.65-1.47) | 0.97 (0.64-1.48) | |
| ≥55 | 1-1 | 251 (74.3) | 195 (74.4) | 1.0 | 1.0 | 119 (77.3) | 1.0 | 1.0 |
| 1-2 or 2-1 | 11 (3.3) | 1 (0.4) | 0.12 (0.02-0.91) | 0.11 (0.01-0.91) | 4 (2.6) | 0.77 (0.24-2.46) | 0.90 (0.26-3.07) | |
| 2-2 | 76 (22.5) | 66 (25.2) | 1.12 (0.77- 1.63) | 1.08 (0.72-1.62) | 31 (20.3) | 0.86 (0.54-1.37) | 0.77 (0.47-1.26) | |
| Smoking | ||||||||
| Nonsmoker | 1-1 | 219 (75.0) | 73 (68.9) | 1.0 | 1.0 | 143 (73.7) | 1.0 | 1.0 |
| 1-2 or 2-1 | 4 (1.4) | 3 (2.8) | 2.25 (0.49-10.30) | 2.07 (0.43-9.95) | 4 (2.1) | 1.53 (0.38-6.23) | 1.43 (0.32-6.52) | |
| 2-2 | 69 (23.6) | 30 (28.3) | 1.30 (0.79-2.16) | 1.28 (0.76-2.16) | 47 (24.2) | 1.04 (0.68-1.60) | 1.11 (0.70-1.74) | |
| Smoker | 1-1 | 271 (74.59) | 233 (71.5) | 1.0 | 1.0 | 154 (78.6) | 1.0 | 1.0 |
| 1-2 or 2-1 | 14 (3.9) | 4 (1.2) | 0.33 (0.11-1.02) | 0.35 (0.11-1.12) | 6 (3.1) | 0.75 (0.28-2.00) | 0.80 (0.29-2.23) | |
| 2-2 | 79 (21.7) | 89 (27.3) | 1.31 (0.92-1.86) | 1.35 (0.94-1.94) | 36 (18.4) | 0.80 (0.52-1.25) | 0.79 (0.51-1.25) | |
| Alcohol consumption | ||||||||
| Nondrinker | 1-1 | 158 (76.0) | 97 (74.6) | 1.0 | 1.0 | 117 (77.0) | 1.0 | 1.0 |
| 1-2 or 2-1 | 3 (1.4) | 2 (1.5) | 1.09 (0.18-6.62) | 1.48 (0.23-9.54) | 2 (1.3) | 0.90 (0.15-5.47) | 1.01 (0.15-6.70) | |
| 2-2 | 47 (22.6) | 31 (23.9) | 1.07 (0.64-1.81) | 1.16 (0.67-2.01) | 33 (21.7) | 0.95 (0.57-1.57) | 1.02 (0.59-1.74) | |
| Drinker | 1-1 | 332 (74.1) | 209 (69.2) | 1.0 | 1.0 | 178 (75.4) | 1.0 | 1.0 |
| 1-2 or 2-1 | 15 (3.4) | 5 (1.7) | 0.53 (0.19-1.48) | 0.58 (0.20-1.69) | 8 (3.4) | 1.00 (0.41-2.39) | 1.25 (0.49-3.19) | |
| 2-2 | 101 (22.5) | 88 (29.1) | 1.38 (0.99-1.93) | 1.45 (1.01-2.07) | 50 (21.2) | 0.92 (0.63-1.36) | 0.86 (0.58-1.29) | |
| Drinking water source | ||||||||
| Tap water | 1-1 | 160 (76.2) | 46 (65.7) | 1.0 | 1.0 | 62 (73.8) | 1.0 | 1.0 |
| 1-2 or 2-1 | 5 (2.4) | 2 (2.9) | 1.39 (0.26-7.41) | 1.33 (0.25-7.21) | 3 (3.6) | 1.55 (0.36-6.68) | 1.77 (0.37-8.49) | |
| 2-2 | 45 (21.4) | 22 (31.4) | 1.70 (0.93-3.12) | 1.77 (0.94-3.32) | 19 (22.6) | 1.09 (0.59-2.01) | 1.13 (0.59-2.16) | |
| Ground water | 1-1 | 137 (70.6) | 106 (71.6) | 1.0 | 1.0 | 97 (82.2) | 1.0 | 1.0 |
| 1-2 or 2-1 | 7 (3.6) | 2 (1.4) | 0.37 (0.08-1.81) | 0.46 (0.09-2.37) | 3 (2.5) | 0.61 (0.15-1.40) | 1.13 (0.26-5.01) | |
| 2-2 | 50 (25.8) | 40 (27.0) | 1.03 (0.64-1.68) | 1.23 (0.72-2.11) | 18 (15.3) | 0.51 (0.28-0.93) | 0.59 (0.31-1.10) | |
| Well water | 1-1 | 162 (77.9) | 123 (74.1) | 1.0 | 1.0 | 93 (71.5) | 1.0 | 1.0 |
| 1-2 or 2-1 | 4 (1.9) | 3 (1.8) | 0.99 (0.22- 4.50) | 1.10 (0.23-5.26) | 1 (0.8) | 0.44 (0.05-3.95) | 0.54 (0.06-4.99) | |
| 2-2 | 42 (20.2) | 40 (24.1) | 1.25 (0.77-2.05) | 1.28 (0.76-2.16) | 36 (27.7) | 1.49 (0.89-2.49) | 1.41 (0.83-2.39) | |
| Natural/ | 1-1 | 31 (70.5) | 27 (64.3) | 1.0 | 1.0 | 41 (75.9) | 1.0 | 1.0 |
| mineral water | 1-2 or 2-1 | 2 (4.6) | 0 (0) | - | - | 3 (5.6) | 1.13 (0.18-7.21) | 0.89 (0.08-9.73) |
| 2-2 | 11 (25.0) | 15 (35.7) | 1.57 (0.62-3.98) | 1.69 (0.59-4.80) | 10 (18.5) | 0.69 (0.26-1.82) | 0.64 (0.21-1.95) | |
CI, confidence interval; OR, odds ratio. a)Polytomous logistic regression was used to assess the association by histological type of gastric cancer, b)Composed of 2 polymorphisms of TLR4 rs10759932 and rs10983755; "1" indicates the major allele of each polymorphism and "2" indicates the minor allele of each polymorphism, c)Adjusted for age, sex, smoking, alcohol consumption, education, and drinking water source during childhood.*p<0.05..