Introduction
The adenomatous polyposis coli (APC) gene, a tumor suppressor gene, encodes a 310 kDa protein which is composed of 2,846 amino acids and is mutated not only in familial adenomatous polyposis but also in most sporadic colorectal tumors [1,2]. These APC mutations are primarily protein truncating, deleting both central domains that regulate Wnt signaling and C-terminal domains that interact with the cytoskeleton. Recent results underscore the clinical utility of APC genetic analysis in the diagnosis, prognosis, and appropriate therapeutic intervention of colorectal cancer (CRC) [3–5]. Therefore, a better description and classification of APC gene mutations would be helpful.
Several studies have shown that the N- and C-terminus of APC have different biological functions, like N-terminal fragments of APC have been shown to cause dominant defects in directional cell migration in multiple model systems [6] and C-terminally truncated proteins present in CRC lack the domains that are required for binding to microtubules, EB1, and β-catenin, potentially leading to the induction of chromosomal instability, activation of proliferation, and inhibition of differentiation [7]. In clinical, Mondaca et al. [8] have revealed that metastatic CRCs (mCRCs) with N-terminal mutation of APC had significantly longer overall survival (OS) and progression-free survival (PFS) versus those with C-terminal side or no APC alteration. The underlying mechanism for these differences is that C-terminal side APC alterations had more concurrent activating mitogenic alterations than N-terminal side APC alterations. In another study, alteration of APC induces programmed death-ligand 1 expression via a β-catenin/TCF4 pathway, then promotes tumor immune evasion [9]. However, in a mice study, the C-terminus of APC does not influence intestinal adenoma development or progression [10]. However, the functional significance of APC specific alterations in Asian mCRC populations is still unclear.
To further study the function of APC specific mutations in mCRC, we explored the association between the tumor mutational burden (TMB) and APC C-terminal or N-terminal mutations in 278 Chinese mCRC patients by a 639 genes sequencing assay, a high throughput, targeted-DNA-sequ-encing panel. We then investigated the co-mutation patterns at the pathway and gene level to understand the underlying mechanism of outcome difference between APC C-terminal and N-terminal alteration. Our research provides new insights into APC specific mutations and a potential new prognostic stratification of patients with mCRC.
Materials and Methods
1. Patients and sample characteristics
From December 2021 to August 2022 at Shanghai Second Affiliated Hospital of Naval Medical University, 275 patients with pathologically diagnosed mCRC were enrolled in the present study. Each patient was pathologically diagnosed and required to obtain the tumor tissue and paired blood sample. Exclusion criteria include that no pathologically confirmed mCRC, no tissue or blood samples were provided, and the cell blocks of samples contained tumor cells less than 20%. Data on clinical characteristics, including age and gender, were retrieved from medical records. Written informed consent was obtained from all participants, and this study was approved by the institutional review board of our hospital (20211218).
2. DNA extraction and library construction
According to the manufacturer’s protocols, the tumor DNA and blood genome DNA were extracted using human tissue DNA extraction kit (Yunying Medicine, Zhejiang, China) and human blood genome DNA extraction kit (Yunying Medicine), respectively. DNA was eluted in the elution buffer, and concentration and purity were assessed using a NanoDrop spectrophotometer. DNA was stored at −20°C until use. Library preparation was constructed using the VAHTS Universal DNA Library Prep Kit for Illumina (San Diego, CA). Target enrichment was performed using Shanghai YunYing’s optimized probes, which target the exons and some introns of 639 cancer-related genes. Sequencing was performed on an Illumina NextSeq500 platform using the manufacturer’s protocols.
3. Next-generation sequencing–based assay and bioinformatics analysis
FastQC software (ver. 0.11.2) and customized python script were used to screen FASTQ files, with the adaptor sequences and sequences with Q below 30 removed. Clean reads were mapped to the reference human genome GRCh37/hg19 using BWA (Burrows Wheeler Aligner ver. 0.7.7). Bam files were then realigned and recalled using GATK3.5, which was also applied to detect mutations. Duplicate sequences were removed using Picard MarkDuplicates (ver. 1.35) to reduce potential polymerase chain reaction bias. VarScan (ver. 2.3.2) was used to select single nucleotide variations (SNVs), satisfying the following criteria: depth ≥ 100, reads ≥ 10, and allele frequency ≥ 5% (if hotspot, ≥ 1%). Pindel (ver. 0.2.5b8) was used for insertion or deletion (indel) detection using default parameters, with at least five unique reads.
Compared with matched normal samples, somatic SNVs and insertion and deletion (InDels) of tumors were named and functionally annotated using MuTect v. 1.1.4 and Varscan2 v. 2.3.9 software. The mutations with variant allele frequency ≥ 5% were defined as high-confidence mutations (≥ 1% for hotspots). TMB was calculated using the number of all somatic, coding, base substitution, and indel mutations per megabase including synonymous mutation. The total number of mutations counted was divided by the size of the coding region of the targeted territory (1.1 Mb of coding genome) to calculate the TMB per megabase. Microsatellite instability (MSI) score of all samples were calculated by MSIsensor [11] with default parameters, a software tool for quantifying MSI in genome sequencing data using tumor only, or paired tumor-normal samples. We used 24 microsatellite sites as input files for MSI detection on tumor only pattern. The MSI score was defined as the percentage of unstable microsatellites in all used microsatellites. Each microsatellite site would have at least 20 spanning reads and single-nucleotide mutations.
4. Public database verification
cBioPortal (https://www.cbioportal.org/) was employed for large sample size verification. The detailed clinical baseline, gene sequencing, and prognosis information of 899 patients with mCRC were downloaded, and the duplicate samples were removed. Samples that did not match somatic mutations were also excluded.
5. Statistical analysis
The prevalence and distribution of genomic alterations are visualized via the R package “maftools”. R package “ggplot2” was used to draw the boxplot. The nonparametric Wilcox test was subsequently used to test for significance in the difference of means between the two populations. R package “Survminer” was used to draw the Kaplan-Meier graph with axis units of months, and the log-rank test p-value, risk table, and censoring table are shown.
Results
1. Patient characteristics
A total of 275 patients newly diagnosed with mCRC cancer were enrolled in the present study. The mean age at diagnosis of the study participants was 60.0 years (range, 13 to 88 years; median, 61 years), and no significant difference was observed between males and females (mean, 60.6 vs. 60.4 years; median, 63 vs. 61 years; p=0.96). Other characteristics of the cohort are summarized in Table 1, and details are displayed in S1 Table. All participants successfully finished targeted sequencing, and the completed sequencing strategy included the whole exons and partial introns of 639 genes indicated in S2 Table.
2. Mutation overview
A missense mutation was the most common mutation type, followed by frameshift deletion and nonsense mutations (Fig. 1A). Single nucleotide polymorphisms represented a larger fraction than insertions or deletions in all variant types (Fig. 1B). The C > T transition was a prevailing SNV in mCRCs (Fig. 1C). As shown in Fig. 1D and E, the number of altered bases in each sample and the variant classification summary were counted. In mCRCs, the top 10 mutated genes were TP53 (76%), APC (69%), KRAS (46%), PIK3CA (19%), FBXW7 (16%), SMAD4 (15%), LRP1B (14%), KMT2D (9%), BRAF (9%), and KMT2C (7%) (Fig. 1F) which might play important roles in mCRC biological processes. According to the waterfall plot of the top 20 mutated genes, where the mutation type is denoted by various colors with annotations, frameshift mutations were mostly observed in the C-terminal side group and other or unaltered group (Fig. 1G).
3. The significant difference in TMB between APC-specific mutation groups
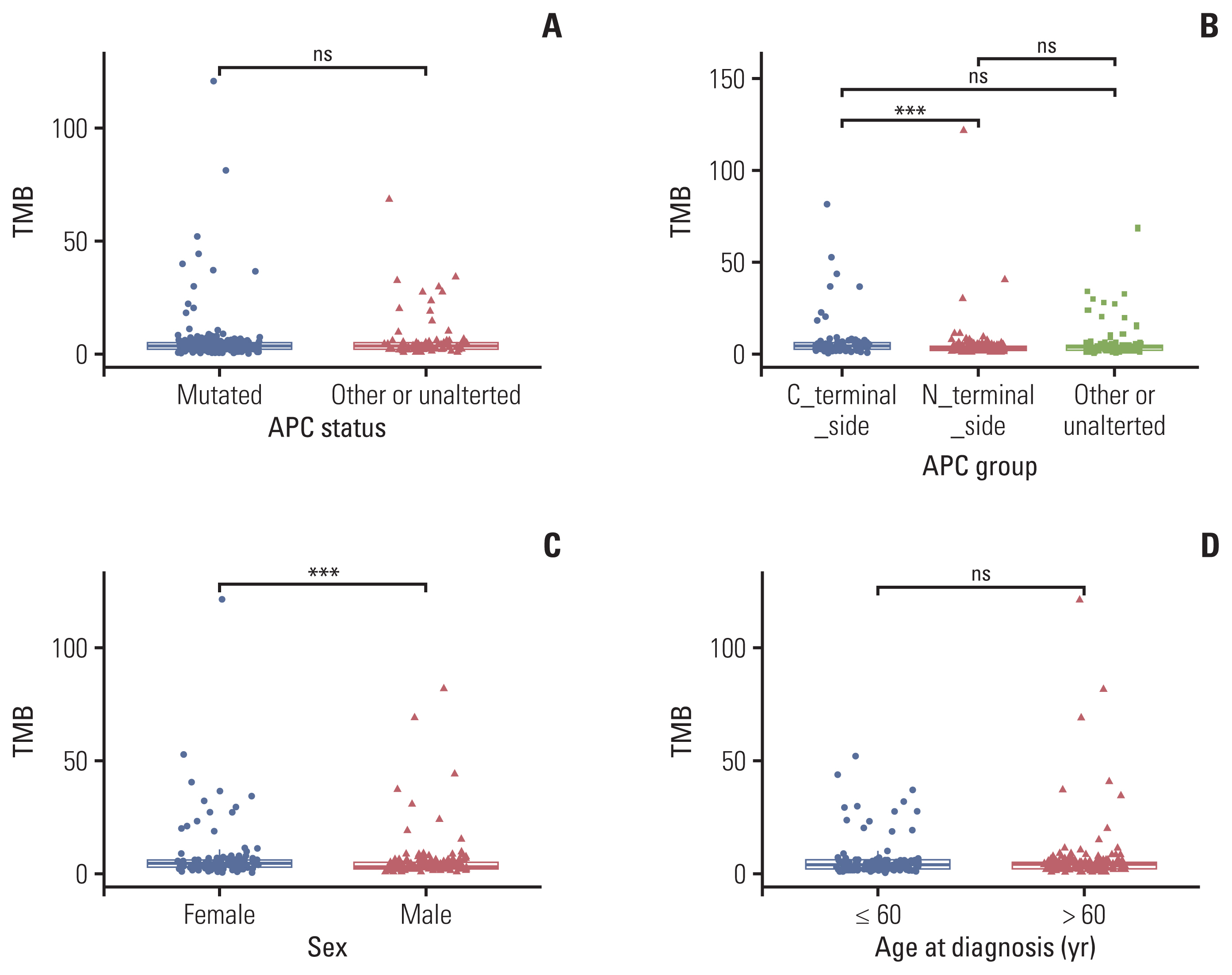
According to the criteria for classifying APC specific mutation groups from a previous study [8], we divided enrolled patients into three groups: APC C-terminal alteration (n=76), APC N-terminal alteration (n=123) and APC unalteration or other mutation (n=76), as Table 1 shown. As Fig. 2A shown, TMB was not significant between APC mutated or not. But TMB was significant between APC C- and N-terminal side alteration (median, 4.4 vs. 2.9 muts/Mb; p < 0.001) (Fig. 2B). TMB also showed a significantly higher score in female patients than in male patients (median, 4.4 vs. 2.2 muts/Mb; p < 0.001) (Fig. 2C). However, TMB showed not any relationship between age at diagnosis group (Fig. 2D). In terms of MSI, no relationship was shown among different APC mutation groups, as well as gender and age at diagnosis (S3A–S3D Fig.) (p > 0.05).
4. Verification of the relationship between clinical indicators and TMB score
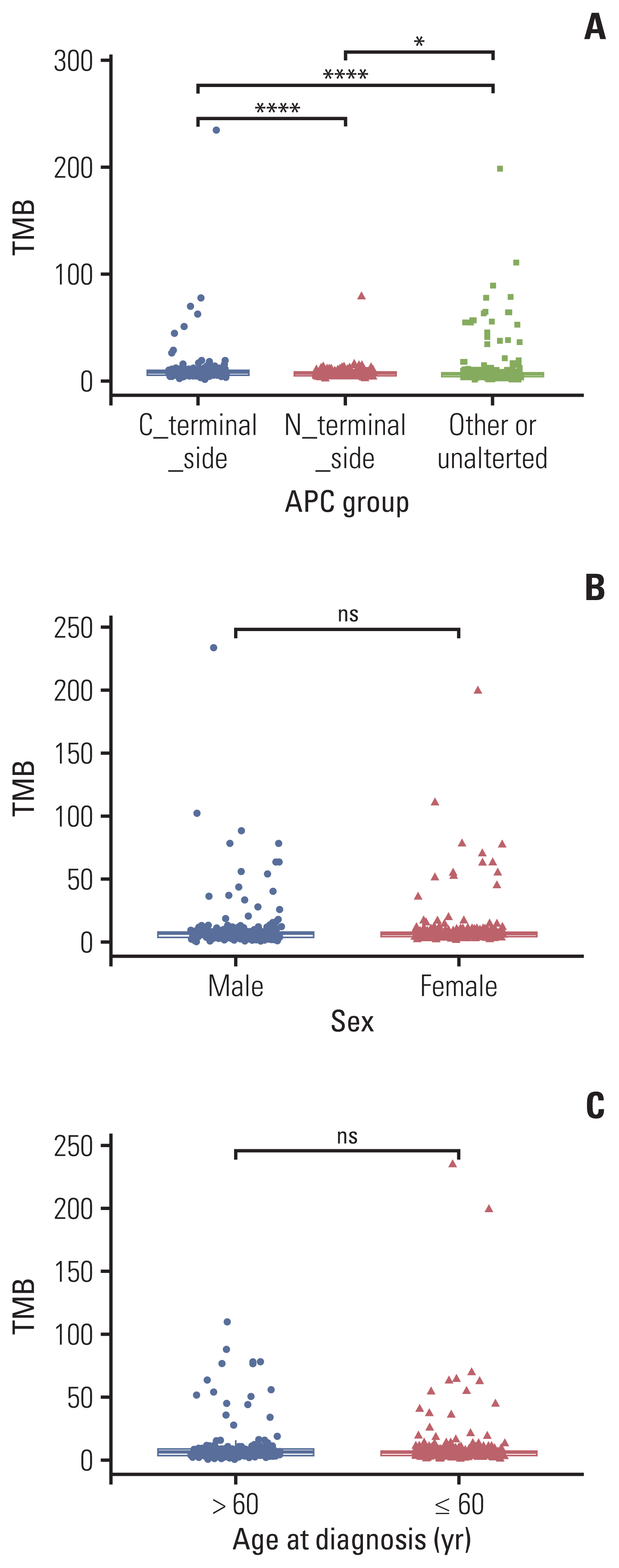
We employed the cBioPortal for verifying our results, which is an open-source resource for the interactive exploration of multidimensional cancer genomics data sets. A total of 899 mCRC patients were enrolled in our verification cohort. We further grouped them by the age at diagnosis in years, gender, or APC mutation status, respectively. The relationship between TMB score or MSI score and diagnosis in years, sex, APC mutation status, and APC specific mutation was analyzed.
In the verification cohort, as Fig. 3A shown, similar tendencies were confirmed that APC C-terminal side mutation was significantly correlated to a higher TMB score than APC N-terminal side mutation (p < 0.001). Similarly, in terms of MSI, there is no significant relationship observed between APC groups and MSI value (S4A and S4B Fig.). Unlike our next-generation sequencing sequencing result, whether TMB or MSI, did not show a significant relationship with sex (Fig. 3B, S4C Fig.), but showed the same trend in the group of age at diagnosis (Fig. 3C, S4D Fig.).
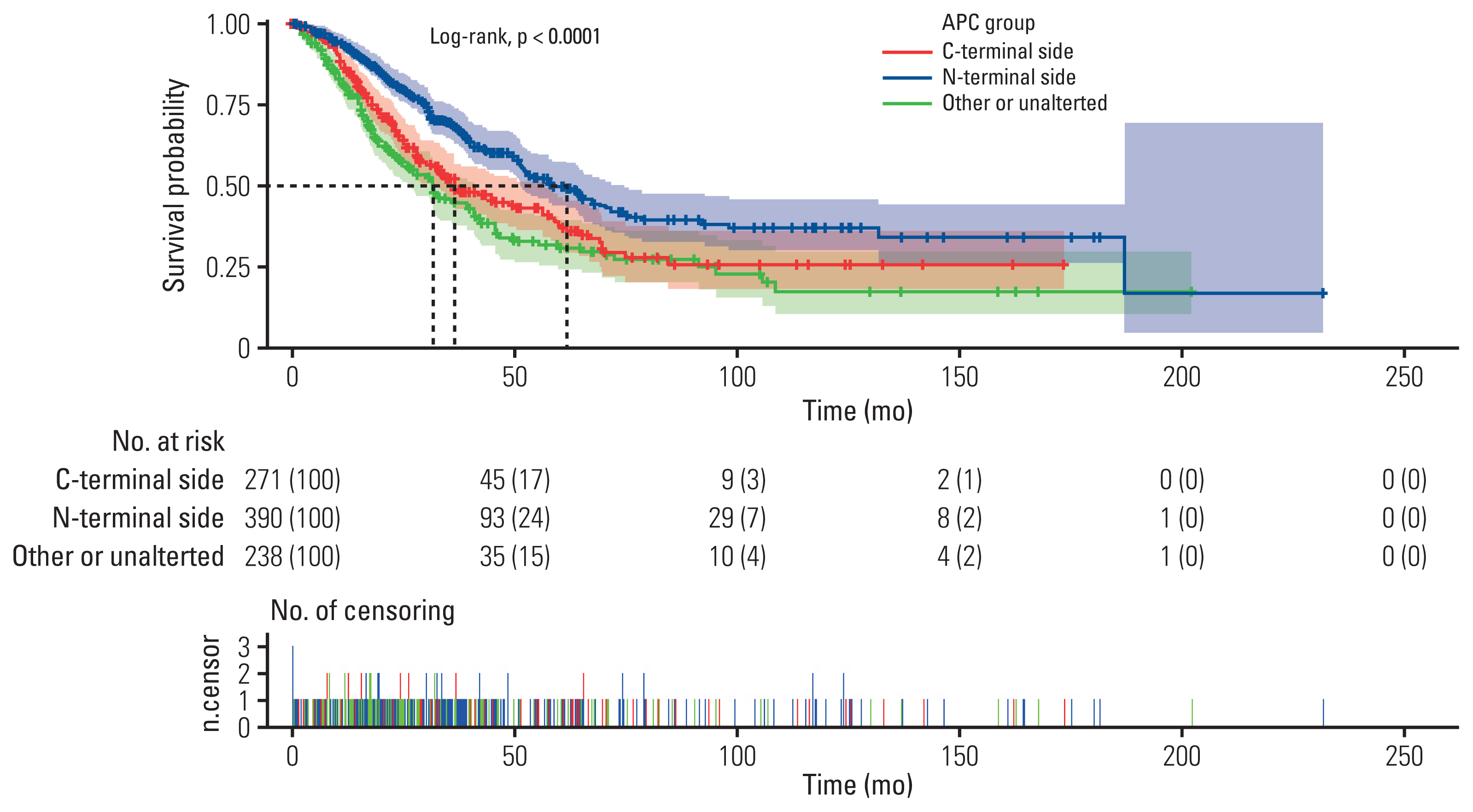
5. Prognostic value analyses
To understand the clinical value of the difference we found, OS for each APC specific mutation was analyzed. As Fig. 4 shown, in metastatic mCRC patients from The Cancer Genome Atlas (TCGA), APC N-terminal side mutation group showed a significantly enhanced OS than the C-terminal side group (61.13 vs. 36.43 months, p < 0.001, n=662), and other or unaltered group (61.13 vs. 31.90 months, p < 0.001, n=628). However, the C-terminal side group showed no significant relationship with the other or unaltered group (36.43 vs. 31.90 months, p=0.072, n=609).
6. Gene-pathway difference analysis
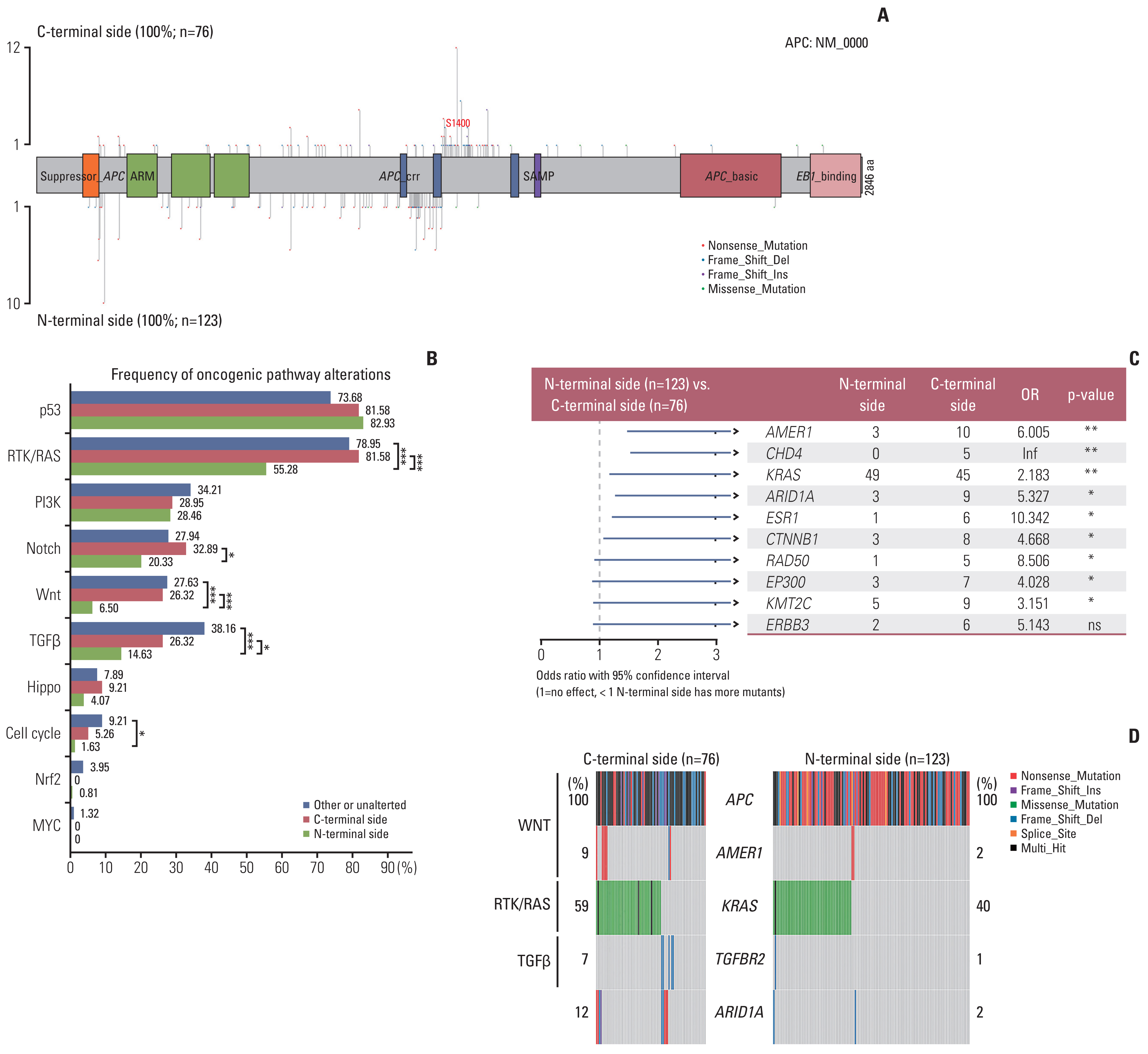
To understand the underlying mechanism for these differences in outcomes, we investigated co-mutation patterns at the pathway and gene levels. According to the results of APC gene mutation distribution, we found mCRC patients with APC C-terminal truncated mutation (nonsense mutation, frame shift deletion, and frame shift insertion) are often accompanied by N-terminal truncated mutation, and these mutations are mainly concentrated near the S1400 position of APC gene (Fig. 5A). Compared with C-terminal side and other or unaltered groups, RAS/RTK, Wnt, and transforming growth factor β (TGFβ) pathway alterations were significantly lower in mCRCs with N-terminal side APC mutations group by analysis for frequency of oncogenic pathway alterations (Fig. 5B). Notch pathway alterations were significantly enriched in mCRCs with N-terminal side APC mutations (p < 0.05). As for the Cell cycle pathway, the N-terminal side group showed a relatively low frequency of alteration especially compared with the other or unaltered group (p < 0.05).
We performed a comparative analysis of the mutation between the N-terminal side group and C-terminal side group, and found that there was a significant difference in the mutation frequency of nine genes (AMER1, CHD4, KRAS, ARID1A, ESR1, CTNNB1, RAD50, EP300, and KMT2C) (p < 0.05) (Fig. 5C).
Actionable mutations analysis was based on Oncology Knowledge Base (OncoKB, www.oncokb.org) database. Overall, four genes in three pathways were identified with significant differences between the N-terminal side group and the C-terminal side group (Fig. 5D). AMER1 mutations were more frequent in tumors with C-terminal side APC mutations (9% vs. 2%). KRAS and TGFBR2 alterations occur-red in 40% and 1% of the N-terminal side, respectively, vs. 59% and 7% of C-terminal side APC mutant cases, respectively. And ARID1A mutations were more frequent in tumors with C-terminal side APC mutations (12% vs. 2%).
Discussion
Previous research split the APC gene truncating mutations into N-terminal and C-terminal sides at amino acid (aa) 1400 where the N-terminal group contains up to two 20R β-catenin–binding repeats, and showed that in patients with mCRC, the N-terminal side APC mutations group had significantly longer OS and PFS than those with C-terminal side or no APC alteration [8]. Therefore, in our study, we divided 275 enrolled mCRCs into three groups: (1) N-terminal side, (2) C-terminal side and (3) other or unalterted (mutation with no effect on APC function or wide type).
For the first time, we found that mCRC patients with C-terminal side APC mutations had significantly higher TMB scores than N-terminal side, which was further confirmed by public databases. TMB is broadly defined as the number of somatic mutations per megabase of interrogated genomic sequence and is believed to be a key driver in the generation of immunogenic neopeptides displayed on major histocompatibility complexes on the tumor cell surface. Compared with the C-terminal side group, the TMB scores of the N-terminal side group is relatively low, so there may be fewer immunogenic new peptides produced on the surface of tumor cells, making it difficult for tumor cells to escape immune system regulation. This may explain why patients with N-terminal side APC mutations have a better prognosis than C-terminal.
Loss of function mutations was more observed in the APC C-terminal side at nearby of the S1400 site than in N-terminal, which may show more effect on APC-related pathway activity. It is the decreased β-catenin binding capacity of mutated APC lacking the C-terminus and therefore increases in Wnt signaling [10]. Interestingly, in addition to the significant differences in RTK/RAS signaling pathways observed between the APC two groups, our results also revealed for the first-time significant differences in Wnt and TGFβ signaling pathways and related gene mutations. Studies have shown that Wnt and TGFβ signaling pathways are related to tumorigenesis and progression in CRC [12–14], and previous research showed that TGFβ could improve CRC migration in vitro [15]. This result shows that compared with unaltered or other APC mutation group, APC N-terminal mutation may inhibit tumorigenesis and progression by Wnt or TGFβ signaling pathway. Additionally, as the previous review of the role of Wnt in tumors [16], deregulated Wnt signaling in the tumor microenvironment supports malignant transformation and disease progression via a variety of mechanisms, such as improved fitness of the cancer stem cell compartment, evasion of tumor-targeting immune responses, and so on. In the late stage of cancer, TGFβ has some positive roles in promoting tumors, like fueling angiogenesis [17], immune escape and metastasis [15]. KRAS, AMER1, TGFBR2, and ARID1A alteration was more frequent in APC C-terminal side than in the N-terminal side and KRAS is also the major effector in APC groups. Patients with KRAS-mutant CRC have a poorer prognosis than those with KRAS-wild CRC, especially in the mCRC [18,19]. This is also consistent with the results of previous clinical studies on the relationship between RAS mutation and the prognosis of CRC [20]. In Wilms tumor, AMER1 (also known as FAM123B or WTX) is a negative regulator of the Wnt/β-catenin pathway by promoting β-catenin ubiquitination and degradation [21]. But, lack of related research in CRC, even though the TCGA consortium reported AMER1 as a frequently mutated gene in CRC [13]. TGFBR2 alterations are almost frameshift deletion, which may cause loss of function. Studies have shown that downregulating or knockdown can promote CRC cell migration and invasion [22,23]. Also, ARID1A mutations are almost frameshift deletions. Wei et al. [24] observed that ARID1A loss was related to the higher TNM stage (p=0.020) and poor pathological classification (p=0.035), largely consistent with Ye et al.’s findings [25]. These previous studies about the most frequent genes in APC C-terminal side mutation may also seem to imply its worse prognosis than N-terminal side ones.
Notably, there are several limitations to this study. First of all, this is a retrospective study, prospective multicenter studies with a larger cohort are needed to gain a better under-standing of the relationship between APC specific mutations and prognoses or immunotherapy benefits. Second, in this study, tumor tissue samples were used for somatic mutations detection, however, tissue biopsy samples depict only a single time frame from a single site, and might be inadequate to characterize a tumor because of intra-tumoral and inter-metastatic heterogeneity, especially in highly heterogeneous CRC. Liquid biopsy, such as circulating tumor DNA enables real-time dynamic detection of tumors, overcomes tumor heterogeneity, and may provide a more comprehensive molecular characterization in CRC. Third, our mechanism hypothesis requires relevant cell and molecular tests to verify the accuracy.
In conclusion, our study developed a potentially novel approach to mCRC patient prognosis stratification based on APC specific mutation. Furthermore, we investigated the characterization of molecular alterations among N-terminal side APC mutations, C-terminal side APC mutations, and other or APC unaltered, and firstly discovered the TMB score, and frequency of RTK/RAS, Wnt, and TGFβ pathway alterations were significantly lower in Chinese mCRC patients with N-terminal side APC mutations than C-terminal side. These results give a new insight to understand the underlying mechanism of APC specific mutation with different prognostic value. This study will serve as a foundation for further molecular mechanism studies to verify the significance of APC specific mutation in mCRC.